Uno degli amminoacidi essenziali per l'uomo, e per gli animali
superiori in genere, è il triptofano. La biosintesi di questa molecola avviene in
microrganismi e vegetali superiori a partire da intermedi della glicolisi e della via dei
pentoso fosfati. L'ultimo passaggio di questo processo metabolico, consistente nella
reazione tra indolo-3-glicerolo-fosfato e L-serina, è catalizzato dalla triptofano
sintasi.
L'enzima è stato isolato da diverse specie di batteri, alghe e funghi; in particolare
mutanti di Escherichia coli e di Salmonella typhimurium in grado di produrre
elevate quantità di triptofano sintasi ne costituiscono una ottima fonte.
Le molecole estratte da questi ultimi due batteri sono quelle principalmente studiate.
Analogamente a quelle di molti altri organismi, sono degli eteropolimeri costituiti da due
subunità a e due subunità b.
Le catene a isolate catalizzano la scissione
dell'indolo-3-glicerolo-fosfato, indicata come reazione a:
b esistono
generalmente come dimeri b2 e
catalizzano la condensazione di indolo e L-serina a L-triptofano, denominata reazione b:
indolo + L-serina
L-triptofano + H2O
Quest'ultimo passaggio richiede la partecipazione del piridossal
fosfato (PLP) come coenzima.
Il complesso bifunzionale a2b2 catalizza
la reazione fisiologica:
indolo-3-glicerolo-fosfato + L-serina
L-triptofano +
D-gliceraldeide-3-fosfato + H2O
L'associazione delle subunità
a
e b nel tetramero determina un aumento delle
velocità delle reazioni parziali rispettivamente di 100 e 30 volte e una affinità per i
substrati maggiore di 1 o 2 ordini di grandezza [61]. A migliorare ulteriormente
l'efficienza catalitica del complesso a2b2
concorre il fatto che l'indolo, prodotto dalla reazione a,
non sembra diffondere nell'ambiente, ma è convogliato direttamente verso il sito attivo b per diventare substrato della reazione b. Secondo una ipotesi supportata da dati strutturali, il
passaggio avviene attraverso un canale intramolecolare che collega i siti attivi [22].
Le influenze reciproche tra le subunità comprendono anche regolazioni di tipo allosterico
che conseguono all'interazione dell'enzima con substrati od altri leganti [61, 62].
3.2 Struttura
La triptofano sintasi da Escherichia coli è quella sulla quale
è stato condotto il maggior numero di studi funzionali. La forma cristallina che ne è
stata ottenuta non è però adeguata a studi strutturali mediante diffrazione di raggi X.
Cristalli del tetramero
a2b2
utili a questo scopo sono invece stati prodotti utilizzando l'enzima da Salmonella
typhimurium [44]. La struttura tridimensionale del complesso è stata determinata con
una risoluzione di 2.5 Å.
Le quattro catene polipeptidiche appaiono disposte quasi linearmente nella sequenza abba a formare una struttura della lunghezza complessiva di
150 Å. Le subunità a sono pertanto separate e poste
all'estremità del complesso; ognuna interagisce con una sola di quelle b che si trovano al centro del tetramero strettamente associate tra
loro. L'ampia superficie di contatto tra le catene a e b adiacenti è in accordo con le reciproche influenze rilevate da un
punto di vista funzionale; l'associazione di leganti ad una subunità è in grado di
determinare nell'altra variazioni delle affinità per il substrato e delle velocità di
catalisi. Un esempio è costituito dal DL-a-glicerolo-3-fosfato
che, legandosi alla catena a, altera la distribuzione
all'equilibrio degli intermedi della reazione tra L-serina e subunità b.
Questi effetti sono verosimilmente dovuti a variazioni strutturali che si trasmettono da
una zona all'altra del tetramero. Studi fisici e cinetici hanno evidenziato come anche
l'attivazione reciproca che consegue all'associazione dei monomeri a
al dimero b2 sia accompagnata da variazioni
conformazionali.
Determinando la struttura dell'enzima legato all'inibitore competitivo
indolo-propanolo-fosfato è stato localizzato il sito attivo della catena a; questo si trova vicino alla superficie di contatto con la
adiacente subunità b.
Ogni monomero b è costituito da due domini strutturali di
dimensioni analoghe. L'ubicazione del sito catalitico, nel quale è legato il PLP, è
stata individuata all'interfaccia dei due domini, vicino al centro della subunità.
I siti attivi a e b di due subunità
adiacenti distano uno dall'altro 25 - 30 Å e appaiono collegati da un
tunnel intramolecolare che partendo dal sito a si sviluppa
lungo la superficie di contatto dei due domini della catena b
fino al sito catalitico b. Le caratteristiche di idrofobicità
e le dimensioni di questo tunnel ne fanno un passaggio ideale per la migrazione interna
dell'indolo dal sito di formazione a a quello b di utilizzo.
3.3 Reazione
b
La reazione
b è quella più
studiata e meglio chiarita tra le due catalizzate dal complesso tetramerico della
triptofano sintasi [62]. E', dal punto di vista chimico, una b
sostituzione in cui il gruppo OH del carbonio b
della L-serina viene rimpiazzato dall'indolo prodotto dalla reazione a.
La partecipazione del PLP come coenzima ha consentito, grazie alle sue caratteristiche
cromoforiche, l'individuazione spettroscopica di una serie di intermedi coenzima-substrato
che si formano durante il processo. La sequenza di questi intermedi proposta per la
reazione e le relative caratteristiche spettroscopiche sono riportate nello schema 1.
In assenza di substrato il PLP forma una base di Schiff con l'e
-ammino gruppo della lisina 87. Alle forme tautomeriche enolimminica e chetoenamminica di
questo composto vengono attribuite le due bande di assorbimento evidenziate per
l'oloenzima, centrate a 412 e 350 nm.
Quando viene aggiunta L-serina il suo gruppo amminico si addiziona alla base di Schiff
interna formando una gem-diammina, ES I, molto instabile; la successiva espulsione del
residuo lisinico dà origine all'aldimmina esterna ES II, più stabile della specie
precedente. Questo intermedio ha un assorbimento massimo a 422 nm ed è caratterizzato da
una forte fluorescenza con un massimo di emissione a 510 nm. Il passaggio successivo è la
rimozione di un protone dal carbonio a della L-serina con
formazione del chinonoide ES III e la successiva deidrossilazione del carbonio b per dare nuovamente una base di Schiff: l'a-amminoacrilato
ES IV.
Nella reazione della L-serina con il dimero b2 si
accumula come intermedio stabile l'aldimmina ES II, mentre con il complesso a2b2 questa decade
rapidamente formando ES III ed ES IV.
La trasformazione di ES II nei composti successivi comporta verosimilmente delle
variazioni strutturali a livello del sito attivo b, che
risultano termodinamicamente sfavorevoli nel dimero b2
e favorevoli nel complesso a2b2.
Nella reazione tra il tetramero e la L-serina, all'equilibrio si ha uno spettro complesso
dovuto al contributo di più intermedi con un massimo di assorbimento a 350 nm che viene
attribuito ad una forma enolimminica dell'amminoacrilato ES IV.
Oltre che dalla presenza delle subunità a l'equilibrio tra gli
intermedi è influenzato da altri fattori quali il legame di effettori allosterici alle
subunità a, il pH, la temperatura, l'organismo d'origine e lo
stato fisico dell'enzima. Relativamente a quest'ultimo fattore sono stati condotti studi
comparativi [68] che dimostrano, oltre alla competenza catalitica dell'enzima nello stato
cristallino, la comparsa nel cristallo di intermedi enzima-substrato analoghi a quelli che
si formano in soluzione, la conservazione degli effetti modulatori esercitati dai leganti
alla subunità a e la sensibilità al pH. Rispetto alla
reazione condotta in soluzione sono però presenti, in alcuni casi, differenze nella
cinetica e nella distribuzione degli intermedi all'equilibrio attribuibili alle forze di
impacchettamento del reticolo cristallino.
La presenza dell'indolo fa procedere la reazione oltre l'intermedio ES IV verso la
sintesi dell' L-triptofano. L'indolo, probabilmente attivato dall'enzima, porta un attacco
nucleofilo al carbonio b dell'amminoacrilato con formazione di
un intermedio chinonoide ES V poco stabile. Dalla successiva protonazione del carbonio a si ottiene l'amminoacido finale legato al PLP a formare una base di
Schiff. L'attacco a questo composto da parte dell'e-ammino
gruppo della lisina 87 porta alla liberazione dell' L-triptofano e alla riformazione
dell'enzima libero.
3.4 Azione di effettori allosterici, pH e ioni, sulla distribuzione
all'equilibrio degli intermedi della reazione
b
L'influenza sulla reazione
b di
leganti delle subunità a, già da tempo
riconosciuta, e quella del pH, evidenziata recentemente nell'enzima cristallino [68], sono
state analizzate più dettagliatamente in un lavoro svolto con triptofano sintasi da Salmonella
typhimurium in soluzione [5, 67]. In questo studio è stata in particolare valutata la
distribuzione all'equilibrio degli intermedi della reazione tra L-serina e complesso a2b2, in funzione di
pH, temperatura, cationi monovalenti, anione fosfato e la presenza di DL-a-glicerolo-3-fosfato, un analogo del substrato delle subunità
a, l'indolo-3-glicerolo-fosfato
Il rapporto tra le quantità delle due specie predominanti all'equilibrio, aldimmina
esterna ES II (lmax=422 nm) e amminoacrilato ES IV (lmax=350 nm), può essere seguito spettrofotometricamente
e dipende dalla variazione della concentrazione di H+ nel modo indicato in
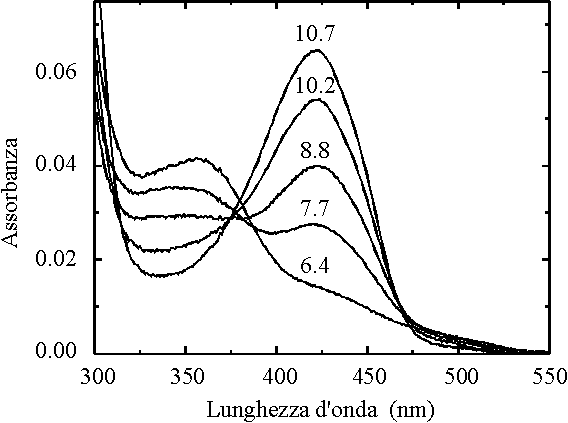
Figura 2.
Figura 2. Spettri di assorbimento della triptofano
sintasi da S. typhimurium in presenza di L-serina a diversi valori di pH
L'aldimmina prevale a pH elevati, mentre al diminuire del pH è sempre
più favorito l'accumulo dell'amminoacrilato. Partendo da una situazione di equilibrio e
variando rapidamente il pH è quindi possibile seguire la ridistribuzione delle molecole
di enzima nelle diverse forme intermedie. La variazione della concentrazione
dell'aldimmina nell'intervallo di pH tra 6 e 10 risulta ben descritta da un curva
corrispondente a quella della ionizzazione di due gruppi con pKa apparenti di
7.6± 0.1 e 10.2± 0.1. Una ipotesi
plausibile è quindi che la trasformazione tra le due forme intermedie dipenda dalla
ionizzazione di due residui amminoacidici direttamente coinvolti nella catalisi. In
alternativa si può pensare che il grado di protonazione dei due residui, che potrebbero
anche non essere localizzate nel sito attivo, influenzino l'equilibrio tra due diverse
conformazioni dell'enzima associate all'aldimmina e all'amminoacrilato.
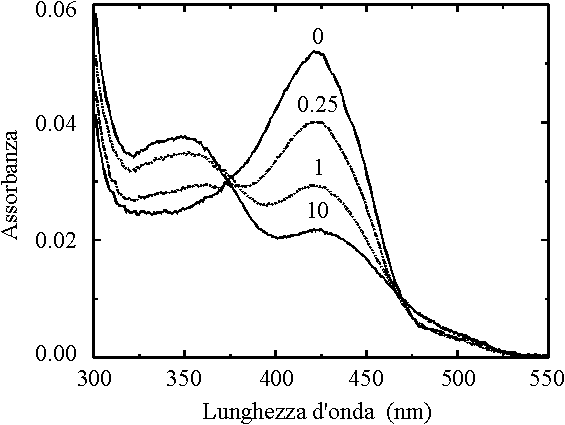
L'effetto del legame del DL-a-glicerolo-3-fosfato alla
subunità a è riportato per l'enzima in soluzione in Figura 3.
Figura 3. Spettri di assorbimento della triptofano
sintasi da S. typhimurium in presenza di L-serina a diverse concentrazioni di
DL-a -glicerolo-3-fosfato (mM)
La presenza di questo composto favorisce nettamente l'accumulo di
amminoacrilato per cui quest'ultimo risulta predominante anche ad alti pH. Il glicerolo
fosfato si lega alla subunità a ad una distanza di circa 25 Å
dal sito attivo b; la sua azione è quindi puramente
allosterica e può consistere nell'induzione di variazioni nella struttura dell'enzima o
nella stabilizzazione di una specifica conformazione, come evidenziato da studi di
luminescenza [87, 88].
Seguendo sia l'assorbimento a 422 nm che la fluorescenza a 500 nm dell'aldimmina ES II è
stato valutato l`effetto di diversi ioni sull'equilibrio aldimmina esterna - a-amminoacrilato. L'anione fosfato esercita un effetto del tutto
analogo a quello del glicerolo fosfato. Per quanto riguarda i cationi si è trovato che lo
ione Na+ stabilizza l'aldimmina esterna, lo ione K+ non sposta
l'equilibrio, mentre gli ioni Li+, Rb+, Cs+ e NH4+
stabilizzano l'amminoacrilato.
Tutti gli effetti osservati in soluzione sono stati riscontrati anche nell'enzima
cristallino.